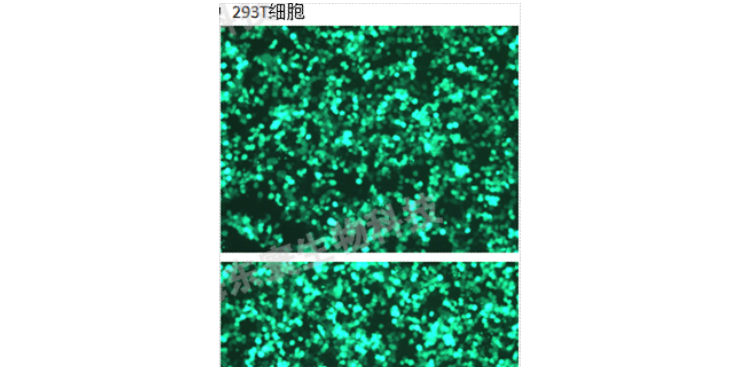
应根据所检测指标的要求,需采取不同策略处理。在如今分子学当道的现实背景下,动物实验除了对实验动物的体征及生理指标进行全的监控外,各种分子检测甚至是组学检测也开始大行其道,动物实验结束之后的机制研究成为了实验的重头戏。一般来说,动物实验检测对象主要包括:(1)体液中各类因子定性及定量检测由于此类检测对象多为蛋白及各类小分子物质,因此在取样过程中应注意防止此类物质降解,在获取后,应及时置于温(≤-80℃)进行保存,在后续实验过程中,也应当注意保存条件的稳定性,避免反复冻融。常用的方法便是酶联免吸附测定(ELISA),除此之外,可利用生化分析仪及不同检测方法的(比色法、比浊法等)方法对各类生化指标及因子进行定性及定量检测。(2)生理变化及免组化检测常用的理检测多使用H&E染色对细胞质及细胞核进行染色标记,以观察细胞变化。除此之外,许多特殊染色也在理生理变化中得到了应用,如利用Masson染色检测纤维化变,Nissl染色检测神经元损伤,β-半乳糖甘酶染色检测细胞衰老等。此外,还可以通过免组化技术对某些特异性标记物进行免显色检测,达到原位定性或定量检测的目的。。细胞污染的处理的技术。北京豚鼠科研技术服务实验室

RNA各种可逆的化学修饰被认为是一种新的表观遗传调控方式。m6A是真核生物mRNA常见的化学修饰,在调控mRNA稳定性,剪切和翻译方面具有重要的作用。作者使用转录组测序发现了METTL3(甲基转移酶3),一种主要的RNAN6-腺苷-甲基转移酶,在人肝细胞(HCC)和多种实体中高表达。在临床上,METTL3的过度表达与肝细胞患者不良预有关。体外实验证明敲除METTL3会抑制HCC细胞增殖,迁移及克隆形成。体内实验证明敲除METTL3会明显抑制HCC体内成瘤和肺转移。另外,使用CRISPR/dCas9-VP64系统,内源性高表达METTL3会促进HCC细胞在体外和体内生长。通过转录组测序、m6A-Seq、MeRIP-PCR,作者确定了SOCS2(细胞因子信号2的抑制因子)作为METTL3介导的m6A修饰的下游靶基因。敲除METTL3表达会消除SOCS2mRNAm6A修饰并增强SOCS2mRNA表达。m6A介导的SOCS2mRNA降解是依赖于m6A“读取器”蛋白YTHDF2。总之,METTL3在HCC大部分高表达中,并通过m6A-YTHDF2依赖机制抑制SOCS2表达从而促进HCC进展。因此,作者发现了在肝发生过程中表观遗传改变的一种新机制。图4RNA甲基化转移酶METLLT3在肝组织中高表达。广西豚鼠科研技术服务购买细胞是生命的基本单位,所有生物体都是由一个或多个细胞组成的。下面就跟着上海东寰一起看看吧。
转录组测序结果及TCGA数据库分析)图5RNA-Seq和m6A-seq联合鉴定SOCS2是介导的m6A修饰的下游靶基因PLoSOne2015,在许多不同种类的RNA中,都已观察到N6-腺苷(m6A)的甲基化,但其在microRNAs中还没有被研究。研究者在FTO1C1,FTO2D4和FTO3C3细胞系中,通过敲除m6A甲基转移酶FTO筛选到表达差异的microRNA,说明miRNA受m6A甲基化的调控。进一步通过MeRIP-Seq发现相当一部分的microRNA具有m6A修饰。通过motif分析,他们发现了区分甲基化和非甲基化microRNA的一致序列。该文章所述的表观遗传修饰在基因表达的转录调控的复杂性上增加了一个新的层次。图FTO敲除对甲基化的miRNAs的稳定状态的影响。参考文献Y,DominissiniD,RechaviG,HeC:Geneexpressionregulationmediatedthroughreversiblem(6)(5):(1):(12):(6):(1):(1):(uridine)(41):(6)(7540):(1):(7481):(4):(6)A-LAIC-seqrevealsthecensusandcomplexityofthem(6)(8):UTRm(6)(4):(7544):(6)(6):(5):(7667):(2):"">panstyle="color:#f5c81c;">xiainducesthebreastcancerstemcellphenotypebyHIF-dependentandALKBH5-mediatedm(6)(14):"">panstyle="color:#f5c81c;">(40):(6)(3):(1):(1):(4):(11):。
转录组和m6A分析显示精子发生相关基因的表达和选择性剪接发生了改变[19]。YTHDC2可促进靶基因的翻译效率,并降低其mRNA的丰度,在精子发生过程中起关键作用。当减数分裂开始时YTHDC2表达上调,YTHDC2敲除小鼠的生殖细胞没有经过偶线期的发育导致小鼠不育[20]。在DNA损伤反应中,METTL3可促进DNA聚合酶κ(Polκ)与核酸剪切修复途径快速定位到UV引起的DNA损伤位点,当缺失METTL3时,细胞无法迅速修复UV照射引起的突变,并且对UV照射更加敏感[25]。在淋巴细胞性小鼠过继转移模型中,Mettl3缺陷通过影响mRNAm6A修饰,降低SOCS家族mRNA衰减,增加mRNA和蛋白表达水平,从而抑制IL-7介导的STAT5活性和T细胞内稳态增殖和分化,进而抑制肠炎的发生[21]。在肝中,METTL14通过调控pri-miRNA的m6A修饰,影响MiR-126的生成加工,从而抑制肝的转移[22]。在乳腺细胞中,低氧刺激能促进依赖低氧诱导因子HIF的ALKBH5的表达,而ALKBH5过表达降低了NANOGmRNA的m6A修饰,从而稳定mRNA提高NANOG的表达水平,终增加乳腺干细胞所占的比例[23]。此外,低氧诱导乳腺细胞中依赖ZNF217的NANOG和KLF4的mRNAm6A甲基化抑制,且ALKBH5敲除降低免疫缺陷小鼠乳腺的肺转移[24]。在肺中。肿瘤细胞侵袭能力检测在学(迁移、侵袭)和免疫学(迁移、趋化)中是常规实验。

是整体甲基化进程所必须的[2]。FTO是ALKB家族的成员,作为个被发现的去甲基酶,可影响剪切因子SRSF2的RNA结合能力,进而调控pre-mRNA的剪切加工过程[3]。目前已发现FTO调节异常与肥胖、大脑畸形和生长迟缓相关,揭示m6A可能对这些疾病具有重要的调节功能[4-6]。ALKBH5是ALKB家族中被发现具有去甲基作用的另一个成员,以RNaseA敏感的方式与核小斑共定位,它可直接催化m6A-甲基化腺苷去除甲基而不同于FTO的氧化去甲基化[7].此外,ALKBH5和它的去甲基化活性影响新生mRNA合的成和剪切效率[7],且ALKBH5敲除雄性小鼠表现出精子发生异常,这可能是精子发生相关基因表达改变的结果[7]。m6AmRNA修饰执行其功能主要通过两个途径:精细调控甲基化转录本的结构,以阻止或诱使蛋白-RNA相互作用;或被直接由m6A结合蛋白识别,诱发续反应。目前一类含有YTH功能结构域的蛋白被鉴定为m6A修饰的结合蛋白。其中YTHDF1,YTHDF2,YTHDF3,YTHDC1和YTHDC2己被证实是m6A的结合蛋白.YTHDF1主要影响m6A修饰基因的翻译,YTHDF2主要影响m6A修饰基因的降解,而YTHDC1结合m6A修饰的基因影响其剪接。HNRNPC是一种丰富的核RNA结合蛋白,参与pre-mRNA的加工[8]。由于大部分研究使用到的是人类来源的细胞,种植到免疫正常的动物体内会发生免疫排斥反应。江苏血液科研技术服务外包
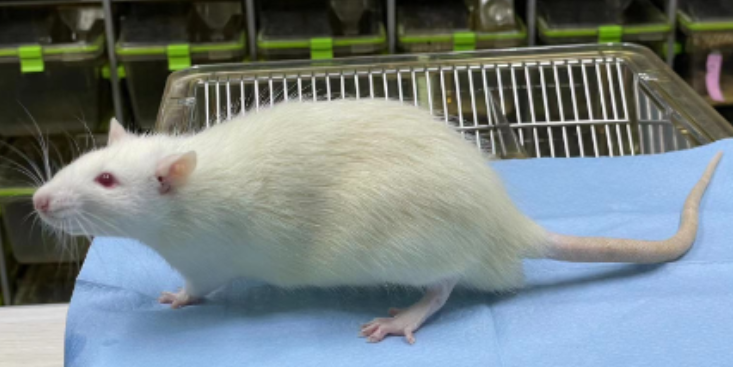
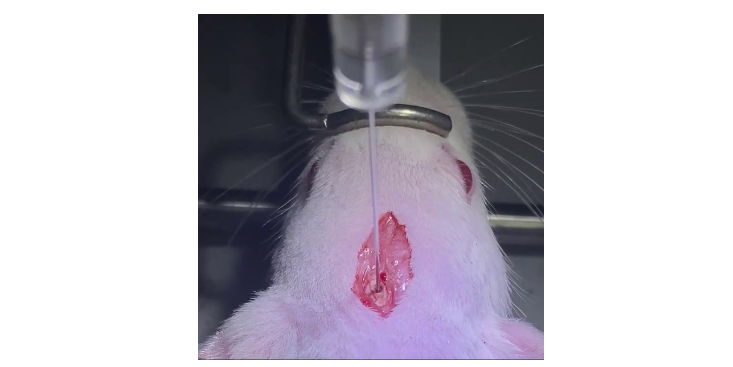
在药物研发过程中,科研人员可以通过对动物模型进行药物处理,观察其疗效和副作用,为新药的试验提供依据。北京豚鼠科研技术服务实验室
动物模型在科研中有着普遍的应用。首先,它们可以帮助科研人员深入理解的共同性,即不同物种之间存在的共有理变化过程。通过对动物模型的研究,科研人员可以更清楚地了解的发展过程和机制,为人类的检查提供理论依据。其次,动物模型还为新研发和苗测试提供了的平台。在研发过程中,科研人员可以通过对动物模型进行处理,观察其和副作用,为新的临床试验提供依据。而在苗测试中,动物模型则可以用来评估苗的性和安全性。此外,动物模型还为科研人员提供了研究人类的跨学科方法。例如,通过比较人类和动物模型的基因组学、蛋白质组学等数据,可以发现与发生相关的关键基因和蛋白质,从而为的和检查提供新的思路。虽然动物模型在科研中发挥了巨大的作用,但也存在一些挑战。首先,由于物种差异的存在,动物模型的表现与人类可能存在差异,因此需要谨慎使用。此外,动物模型的伦理问题也不容忽视,科研人员需要在符合伦理规定的前提下进行相关研究。尽管存在挑战,动物模型的发展前景仍然值得期待。随着科技的不断进步,科研人员将能够开发出更为精确、实用的动物模型。北京豚鼠科研技术服务实验室