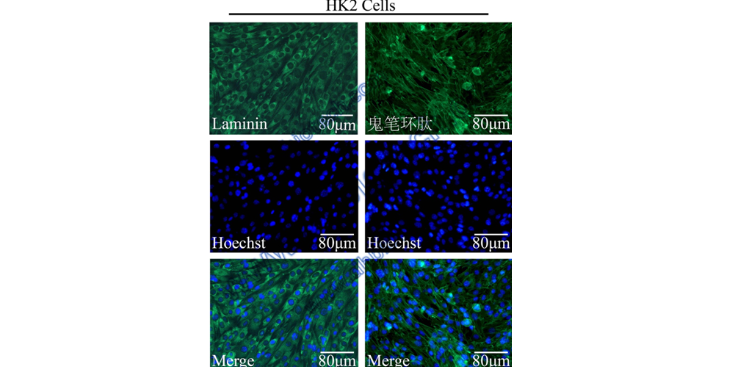
建议按照2×106每孔的数量将293T细胞均匀铺入。(2)第二天:在24小时之内,观察293T细胞的汇合度在90%~95%之间时,向其中加入DNA-脂质体复合体,DNA-脂质体复合体制备方法如下:a)轻轻混匀LipoMax,根据说明书加入相应量于500µlOpti-MEM无血清培养基中,混合均匀并置于室温5分钟。b)在500µlOpti-MEM无血清培养基中稀释DNA,总质量为15µg按照载体质粒:psPAX2:=4:3:1的比例加入DNA。c)将稀释后的LipoMax和稀释后的DNA轻轻混匀,常温静置20分钟,形成DNA-LipoMax复合体。(3)将DNA-LipoMax复合体轻柔地滴加至细胞培养皿中,轻轻摇晃培养皿混匀,放入细胞培养箱中培养。(4)病毒收集浓缩病毒:加入DNA-LipoMax复合体48小时后,收集病毒上清,同时加入10ml预温的293T培养基到细胞培养皿中。将收集到的病毒上清存在4℃冰箱中;收集72小时病毒上清,与48小时病毒上清混在一起。将离心机温度降温到4℃,600g,离心5分钟,去除其中的细胞碎片,上清液经µm滤头过滤,加入病毒浓缩液,配制浓缩病毒液。将浓缩后的病毒放于4℃冰箱摇床上,旋转过夜。第二天,4度离心机,3000~4000g离心15分钟。弃掉上清液,加入1Xpbs或培养基重悬。protocol 分享|过表达细胞株的构建。贵州豚鼠科研技术服务购买
静置25分钟后把酒精倒干,用吸水纸吸出多余的酒精,然后配压缩胶,同样的操作,关键是梳子要插得快,要小心梳子下产生气泡,然后静置30分钟。如果是当天跑胶,我会等上层胶凝2个小时再用,但要注意防干燥缩水,可以在一个小时的时候沿着梳子上缘加点电泳液。所以我一般提前一晚制胶,泡于纯水或者电泳液里置于4度冰箱暂存。三、蛋白电泳1、上样前准备把胶组装到电泳芯上,注意密闭性(否则漏液),如果内槽漏液就不是匀强电场了,条带可能就不是一条直线。然后内槽倒满电泳液,拔梳子,这一步要小心,梳子要两边一起缓缓往上拔出,然后观察泳道内有无脱落的胶粒或者胶丝,有的话用1毫升注射器吸出。然后从冰箱取出蛋白样品,解冻。准备振荡器。2、上样和电泳注意,上样后蛋白会开始慢慢在胶中弥散,所以上样越快越好。我习惯先上蛋白Marker,再上蛋白样品,蛋白上样前确保样品完全解冻和充分振荡(推荐使用振荡器振荡),吸的时候没有拉丝即可,建议上样分钟把样品从冰上取出来,不然样品中SDS可能会结晶析出,从而影响电泳效果。上层胶80V25分钟,下层胶120V65分钟。四、转膜1、转膜前准备我会在电泳结束0分钟准备,把转膜液配好置于4度冰箱预冷,然后裁膜,准备转膜装置。贵州豚鼠科研技术服务分离常用实验动物模型按产生原因分为以下5类:自发性动物模型、诱发型模型、遗传工程模型、医学模型阴性模型。
图2MAZTER-Seq实验流程图图3MAZTER-MINE分析m6A示意图接下来作者便是要验证这一新方法的可行性了。在酵母中敲除IME4的情况下,检测到的剪切效率高于野生型(剪切效率高低m6A水平),m6A抗体富集后的样品剪切效率也低于未富集的Input组。整体水平可靠,那检测的特异性位点是否准确呢?作者也将该方法检测到的新甲基化位点使用放射标记层析检测,发现预测的位点准确存在而且与剪切效率相符合。如图5所示。而图6中,作者则是与m6A抗体IP的方法进行了比较,也证实了这一方法的可行性。图5MAZTER-Seq检测结果验证图6MAZTER-Seq与m6A-Seq比较分析此外,后文中作者也在大规模的CRISPR-Cas9改变m6A状态和酵母减数分裂模型中检测了MAZTER-Seq这一系统;并进一步通过这一方法检测了哺乳动物不同细胞间m6A水平的保守性;也探究了去甲基化酶FTO对整体m6A甲基化水平的影响等。这里小编主要给大家分享这一新技术,其他部分暂不过多分析了。新的技术能拓展我们的研究内容;对于这一技术。
m6A修饰图谱构建及作用机制:通过m6A甲基化测序(MeRIP-Seq,miCLIP)构建疾病细胞模型或者发病组织的m6A修饰谱,分析m6A的motif,peaks数量及分布,Peak关联基因的特征,联合RNA-seq研究m6A甲基化与表达的关系。m6A研究思路方案一方案二研究案例1、.(IF=)为研究ALKBH5的m6A作用机制,作者利用芯片和m6A-seq筛选到胶质瘤增殖相关的FOXM1,通过qPCR、WB、免疫荧光、核质分离WB/qPCR、RIP和MeRIP等实验证明ALKBH5通过去甲基化调节FOXM1在GSCs中的表达。为研究ALKBH5对FOXM1的作用是否受其他因子的调节,作者研究了FOXM1的邻近基因,发现lncRNAFOXM1-AS与FOXM1序列互补,且共表达、共定位,进一步通过RIP,RNApulldown等实验证明lncRNAFOXM1-AS促进ALKBH5和FOXM1初级转录本的相互作用。通过细胞实验进一步验证ALKBH5在lncRNAFOXM1-AS的作用下维持FOXM1的表达和细胞增殖,从而维持GSCs的干性。图3ALKBH5敲除细胞中m6A修饰的特征和基因表达的变化2、RNAN6-methyladenosinemethyltransferaseMETTL3promoteslivercancerprogressionHepatology,2017.(IF=)表观遗传改变极大地促进了人类症的发生。传统的表观遗传研究主要集中在DNA甲基化,组蛋白修饰和染色质重构。近。生物分子学是研究生命体系中分子结构、功能和相互作用的学科。

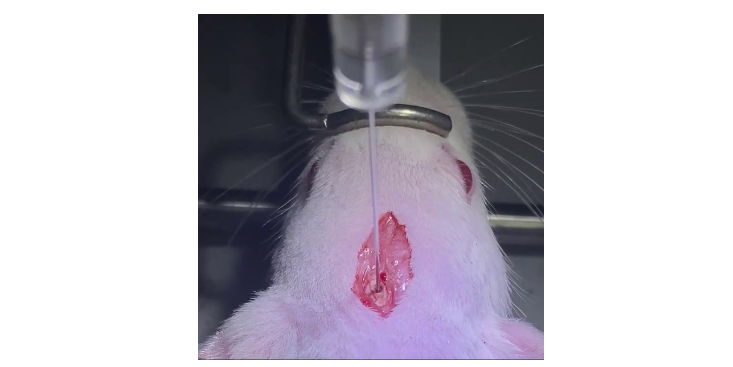
RNA甲基化修饰(m6A)研究RNA甲基化修饰约占所有RNA修饰的60%以上,而N6-甲基腺嘌呤(N6-methyladenosine,m6A)是高等生物mRNA和lncRNAs上为普遍的修饰。目前发现microRNA,circRNA,rRNA,tRNA和snoRNA上都有发生m6A修饰。m6A修饰主要发生在RRACH序列中的腺嘌呤上,其功能由“编码器(Writer)”、“消码器(Eraser)”和“读码器(Reader)”决定[1]。“编码器(Writer)”即甲基转移酶,目前已知这个复合物的成分有METTL3,METTL14,WTAP和KIAA1429;而ALKBH5和FTO作为去甲基酶(消码器)可逆转甲基化;m6A由m6A结合蛋白识别,目前发现m6A结合蛋白(读码器)有YTH结构域蛋白(包括YTHDF1,YTHDF2,YTHDF3,YTHDC1和YTHDC2)和核不均一蛋白HNRNP家族(HNRNPA2B1和HNRNPC)。m6A酶系统METTL3是早先被鉴定为结合SAM的组件,其缺失引起小鼠胚胎干细胞、Hela细胞和HepG2细胞中m6Apeaks的减少。METTL3及其同源蛋白METTL14定位在富含剪切因子的细胞核内亚细胞器-核小斑(Nuclearspeckle)上,显示m6A修饰可能和RNA的剪切加工相关。WTAP与METTL3–METTL14二聚体相互作用,并共定位于核小斑,影响甲基化效率,参与mRNA剪。而KIAA1429作为候选的甲基转移酶复合体的新亚基。总的来说,动物疾病模型是一种重要的研究工具,可以帮助科学家们更好地了解疾病的发生机制和开发新方法.山西模式科研技术服务分离
细胞划痕(wound healing)法是简捷测定细胞迁移运动和修复能力的方法。贵州豚鼠科研技术服务购买
shRNA)蛋白检测蛋白纯化蛋白分析蛋白修饰细胞生物学检测药物筛选实验动物临床检测试剂相关检验试剂抗体库抗体抗体制备第二抗体试剂盒抗体相关抗体库抗体抗体制备第二抗体试剂盒抗体相关技术服务库整体实验外包服务细胞生物学服务测序/分子生物学服务生物芯片服务蛋白相关服务新药研发外包服务技术服务库整体实验外包服务分子生物学服务寡核苷酸合成细胞生物学服务干细胞技术服务微生物学服务免疫学服务蛋白相关服务生物芯片服务实验动物服务新药研发外包服务大型仪器测试与验证服务仪器维修服务技术培训服务其它服务活动专题CellSignalingTechnology学堂生物标志物检测将如何推进抗药物研发细胞焦亡信号通路关键蛋白和研究动态2018免疫与生物网络研讨会期精选课程Webinar直播企业学堂微芯片上的生化室已有244061人观看轻松搞定疫苗的作用机制已有219615人观看Medidata如何改善患者在临床研究中的体验已有214453人观看安捷伦2017二代测序系列讲座之2:靶标富集和文库构建技术大观Merck新型解决方案原位RNA表达检测方案及基础研究实例分析BIO-RAD学堂GE技术大咖秀CellSignalingTechnology学堂技术专题第十一届中国生物产业大会暨第三届“中国光谷”国际生命健康产业博览会火热。贵州豚鼠科研技术服务购买